DNA N6-甲基腺嘌呤(N6-methyladenine, 6mA)是一种在原核生物中广泛存在的表观遗传修饰,但是高等真核生物中6mA的存在一直是一个难解的谜。2015年,汪海林组与陈大华组合作在Cell上发表了关于果蝇DNA 6mA修饰的研究成果。哺乳动物DNA 6mA的研究也随后开启了新的里程。
但是,由于哺乳动物DNA在样品制备和分析过程中,容易受到原核生物DNA污染,而原核生物6mA 水平远高于哺乳动物的,为哺乳动物DNA 6mA的准确检测与研究带来巨大挑战。同时,研究人员也因此困惑:哺乳动物基因组DNA中是否存在复制后可遗传性的N6-腺嘌呤甲基化修饰?该问题一度引起业内广泛关注与争议。
为了回答这一问题,汪海林研究组发展了一套独有的、无污染的6mA分析方法,在样品预处理、超高效液相色谱串联质谱(UHPLC-MS/MS)分析中排除可能的原核生物DNA的污染,可获得准确的6mA信号。利用这一先进的分析方法,在三种人细胞系和小鼠胚胎干细胞(mouse embryonic stem cells, mES细胞中检测到6mA(图1a, b)。研究还发现6mA可在细胞G1期累积。
为了进一步探索6mA的来源,该组采用他们研发的一种独特的稳定同位素标记方法。该法使用[15N5]-dA作为初始示踪剂处理细胞。其中,[15N5]-dA通过补救合成途径以[15N4]-dA的形式掺入基因组DNA中。理论上也可用于识别哺乳动物细胞内甲基化转移酶产生的6mA ([15N4]-6mA)。研究发现,经[15N5]-dA处理后,细胞DNA被[15N4]-dA有效标记,但未能在mESC等细胞系、不同细胞周期中检测到任何[15N4]-6mA。
该研究采用了第二种有效的同位素标记试剂,即[13CD3]-L-甲硫氨酸,但仍然未检检测到任何[13CD3]-6mA。以上结果表明,在哺乳动物细胞系中检测到的6mA并不依赖于甲基化转移酶。
他们推测细胞内6mA来源于特定DNA聚合酶的活动。据此,采用6mA核苷直接处理mES细胞,基因组DNA中可检测到剂量依赖的6mA掺入。随后,分别用同位素标记的[15N5]-6mA和RNA核苷[CD3]-m6A处理细胞进行验证,可分别检测到[15N5]-6mA和[CD3]-6mA掺入。
那么mES细胞6mA是如何进入基因组的呢?研究发现,与非同步化的细胞相比,G1期细胞中DNA聚合酶X家族成员非模板依赖性的Polλ的mRNA表达增加,而Poly λ敲低则显著降低了G1期和sub G1期6mA水平。且Polλ敲除可导致[CD3]-m6A处理掺入的6mA减少。



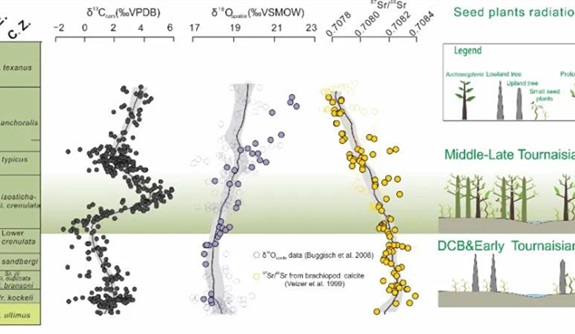
种子植物的辐射可能是触发晚古生代冰期直接原因
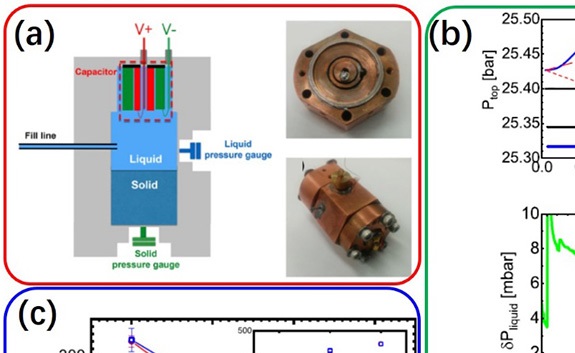
原位测量量子液体中同位素杂质浓度技术